ENO2-MIF轴调控M2巨噬细胞极化,驱动结直肠癌肝转移

文献速览
结直肠癌肝转移(ColorectalLiverMetastasis,CRLM)是患者死亡首要原因,而现有靶向及免疫治疗疗效有限,M2型巨噬细胞介导的免疫抑制微环境是转移发生与治疗耐药的核心诱因,但肿瘤细胞调控巨噬细胞极化的关键机制尚不明确。ENO2作为经典糖酵解酶,已被证实参与肿瘤恶性进展,但其是否介导结直肠癌免疫微环境重塑、调控肝转移进程仍不清楚。本研究依托单细胞与空间转录组,系统解析ENO2在结直肠癌肝转移中的作用,阐明ENO2通过结合稳定MIF、诱导M2巨噬细胞极化的分子机制,并筛选靶向该互作的小分子抑制剂,为结直肠癌肝转移提供新的机制靶点与临床干预方向。

研究路线
- 单细胞图谱解析CRLM细胞特征,筛选转移特异关键基因ENO2;
- 临床队列与生信分析验证ENO2高表达与不良预后、肝转移的相关性;
- 体内外验证ENO2促进结直肠癌恶性进展与肝转移;
- 机制探究:ENO2直接结合MIF,抑制CHIP介导的泛素化降解;
- 阐明ENO2招募HSP90形成复合物,竞争性保护MIF不被降解的分子细节;
- 验证ENO2-MIF轴诱导M2巨噬细胞极化、促进肝转移;
- 筛选吡硫醇为ENO2-MIF互作抑制剂,验证其体内抗肝转移效果。
主要结果
1.单细胞图谱揭示肝转移灶肿瘤细胞恶性特征显著增强
CRLM肝转移灶相比原发灶肿瘤细胞CNV(拷贝数变异)评分更高、基因组不稳定性更强,增殖及缺氧相关基因显著上调;细胞功能层面表现出更强的增殖能力与缺氧适应能力,明确肝转移灶肿瘤细胞具有更高恶性表型。
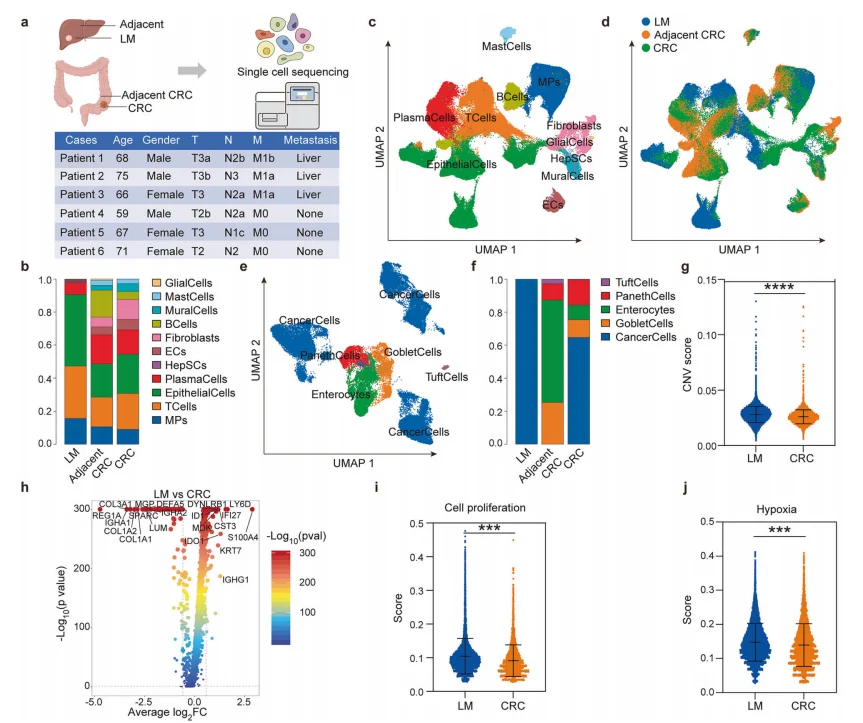
图1.单细胞图谱揭示肝转移病灶中恶性程度增强
2.模块分析筛选出转移原发肿瘤细胞亚群及核心基因ENO2
单细胞模块分析鉴定出具有转移潜能的肿瘤细胞亚群,筛选Module11差异基因,结合预后分析锁定ENO2为核心候选;该亚群高表达EMT特征,是原发灶中具备转移起始能力的细胞群体。
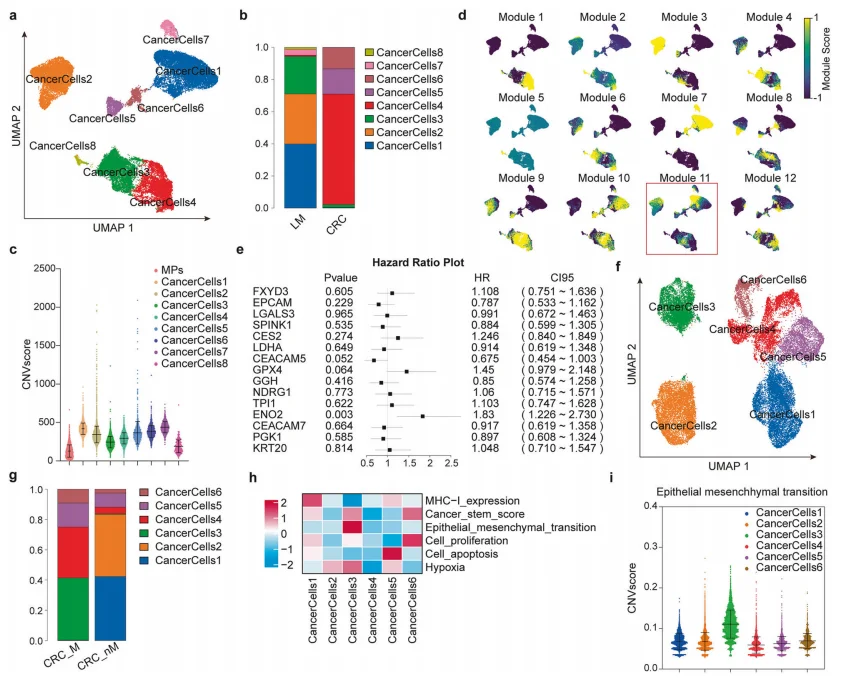
图2.基于模块的转移前癌细胞亚群发现
3.ENO2高表达预示结直肠癌不良预后,且显著促进体内外肝转移
多临床队列及公共数据库验证发现,ENO2在结直肠癌原发灶及肝转移灶显著上调,高表达患者生存期显著缩短;ENO2敲除可抑制肿瘤生长、减少肝转移结节,药物抑制ENO2同样可阻断转移进程,且ENO2可正向调控EMT标志物表达。
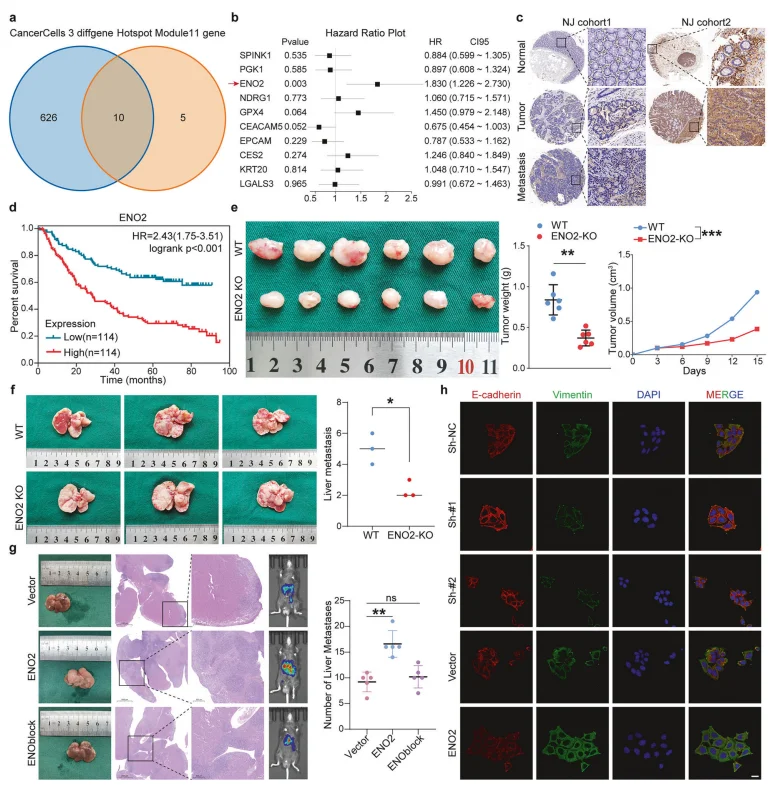
图3.ENO2驱动转移性进展并作为治疗靶点
4.ENO2与MIF直接互作,调控肿瘤-巨噬细胞通讯并诱导M2极化
Cell Chat细胞通讯分析显示ENO2⁺肿瘤细胞MIF信号通路高度活化;Co-IP、GST-pull down及分子突变证实ENO2与MIF直接物理结合;空间转录组证实ENO2⁺肿瘤细胞与M2巨噬细胞在肝转移灶显著共定位,MIF介导肿瘤与巨噬细胞的信号互作,驱动M2极化。
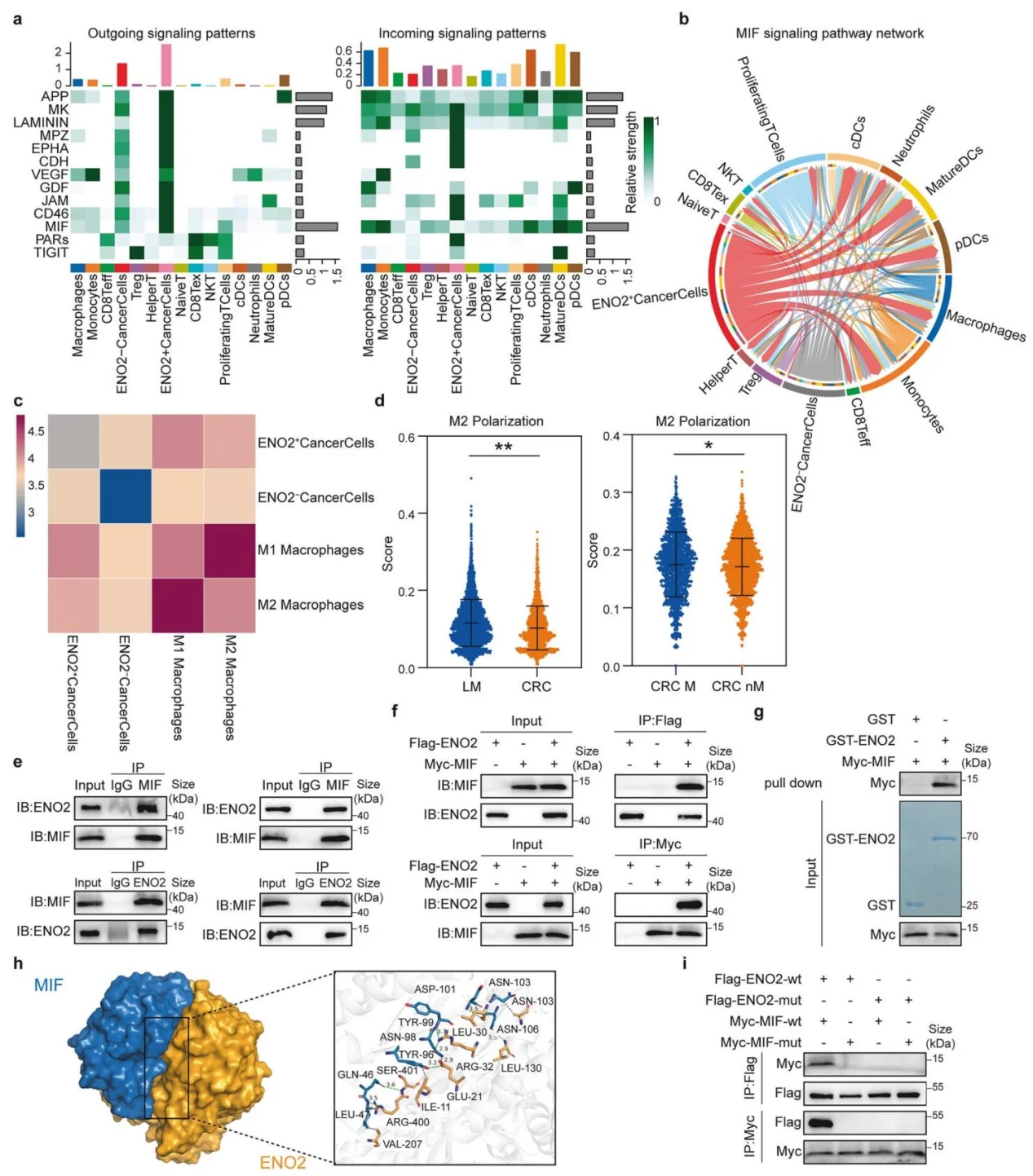
图4.ENO2通过与MIF直接相互作用调控M2型巨噬细胞极化
5.ENO2抑制CHIP介导泛素化降解,稳定MIF并激活下游通路
ENO2可上调MIF蛋白水平,不影响转录;通过抑制E3泛素连接酶CHIP介导的MIF泛素化降解,延长MIF蛋白半衰期;进而激活STAT3、NF-κBp65磷酸化,启动下游促转移及免疫抑制信号。
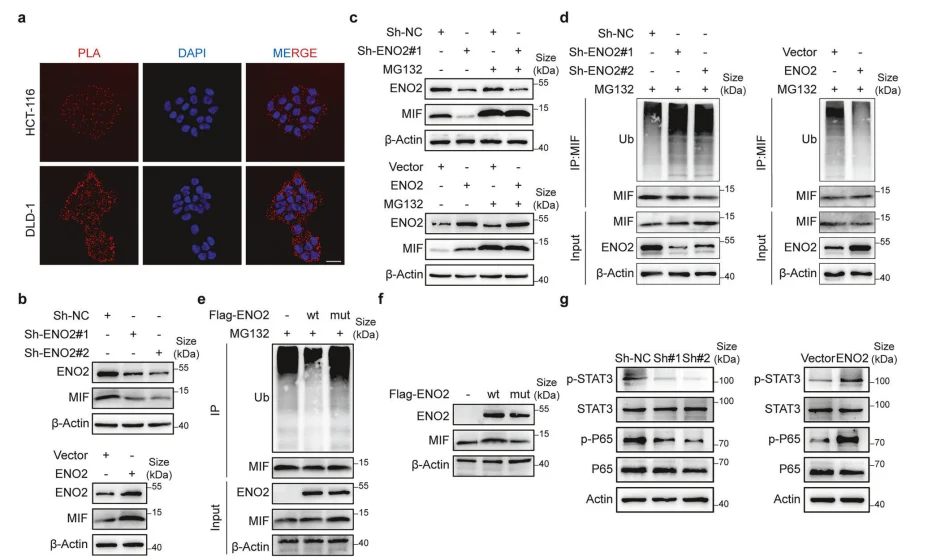
图5.ENO2通过抑制泛素介导的降解来稳定MIF
6.ENO2-MIF轴体内外调控M2巨噬细胞极化并驱动肝转移
细胞共培养、患者类器官共培养模型证实,ENO2过表达上调M2标志物、下调M1标志物;动物实验中MIF抑制剂、M2极化抑制剂可逆转ENO2促转移作用,过表达MIF或M2激动剂可回补ENO2敲除的转移抑制表型,明确ENO2依赖MIF介导M2极化促肝转移。
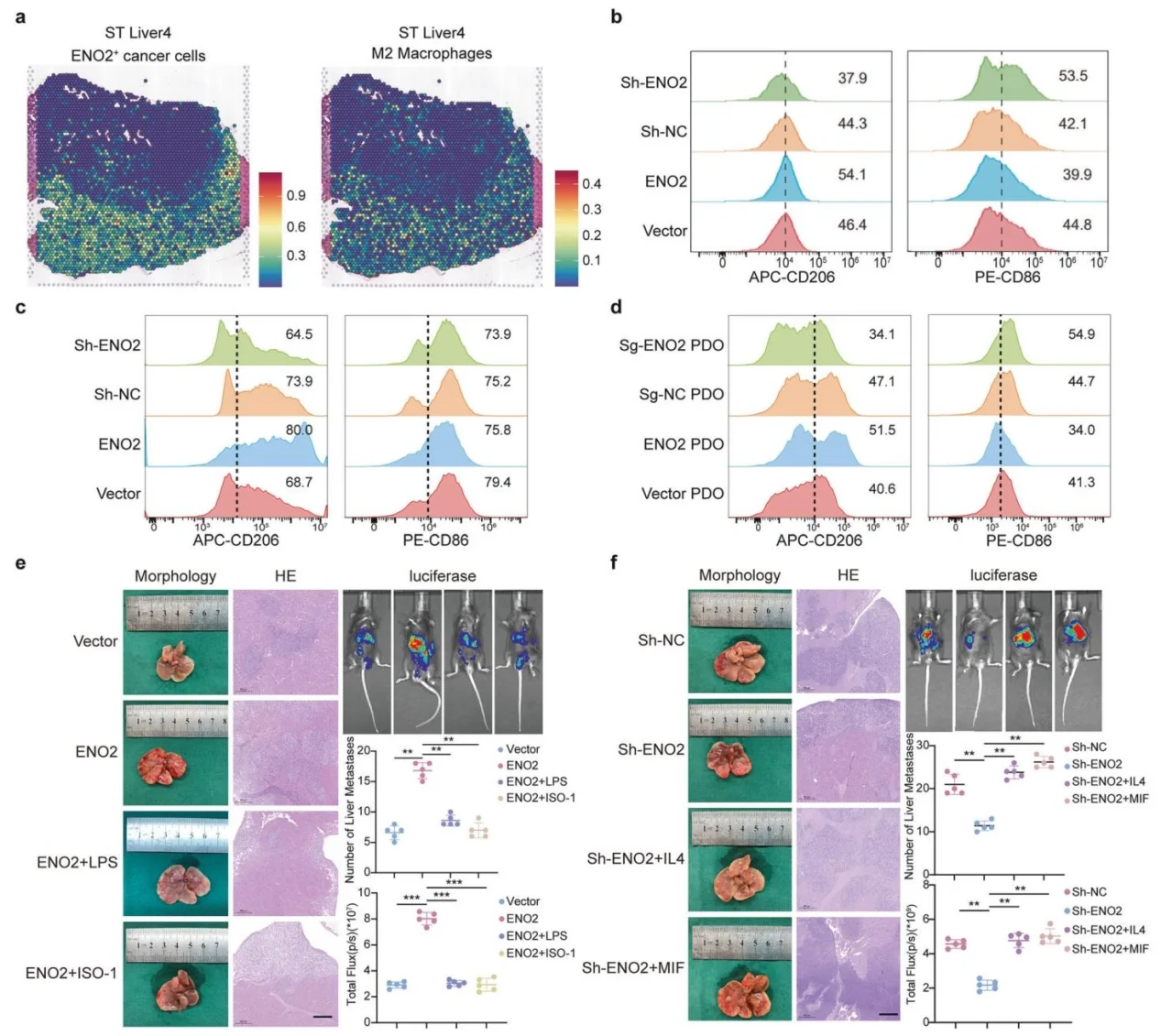
图6.ENO2通过诱导M2型巨噬细胞极化促进肝脏转移
7.ENO2招募HSP90形成三元复合物,拮抗CHIP降解MIF
ENO2可招募分子伴侣HSP90,形成ENO2-HSP90-MIF三元复合物;竞争性阻断CHIP对MIF的识别,锁定MIFK66位为关键泛素化位点,从分子层面阐明ENO2稳定MIF的完整机制。
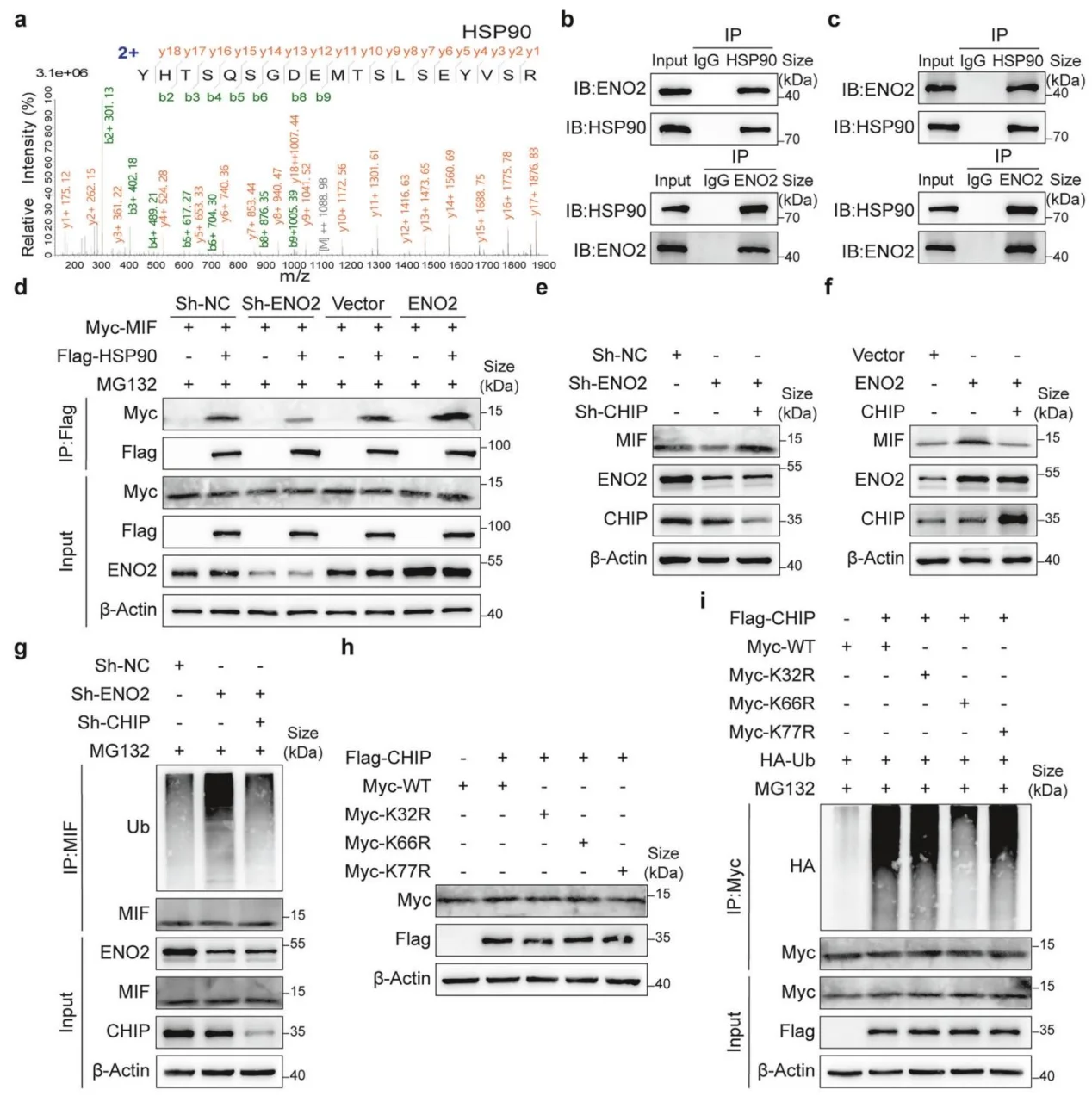
图7.ENO2招募HSP90以拮抗CHIP介导的MIF泛素化及降解过程
8.筛选吡硫醇为ENO2-MIF互作抑制剂,有效抑制结直肠癌肝转移
通过虚拟筛选、分子动力学模拟鉴定吡硫醇(Pyrithioxin)可靶向结合ENO2-MIF互作界面,破坏二者结合、促进MIF泛素化降解;体内给药可显著降低肝转移负荷,下调MIF及下游STAT3、NF-κB活化,具备良好临床转化潜力。
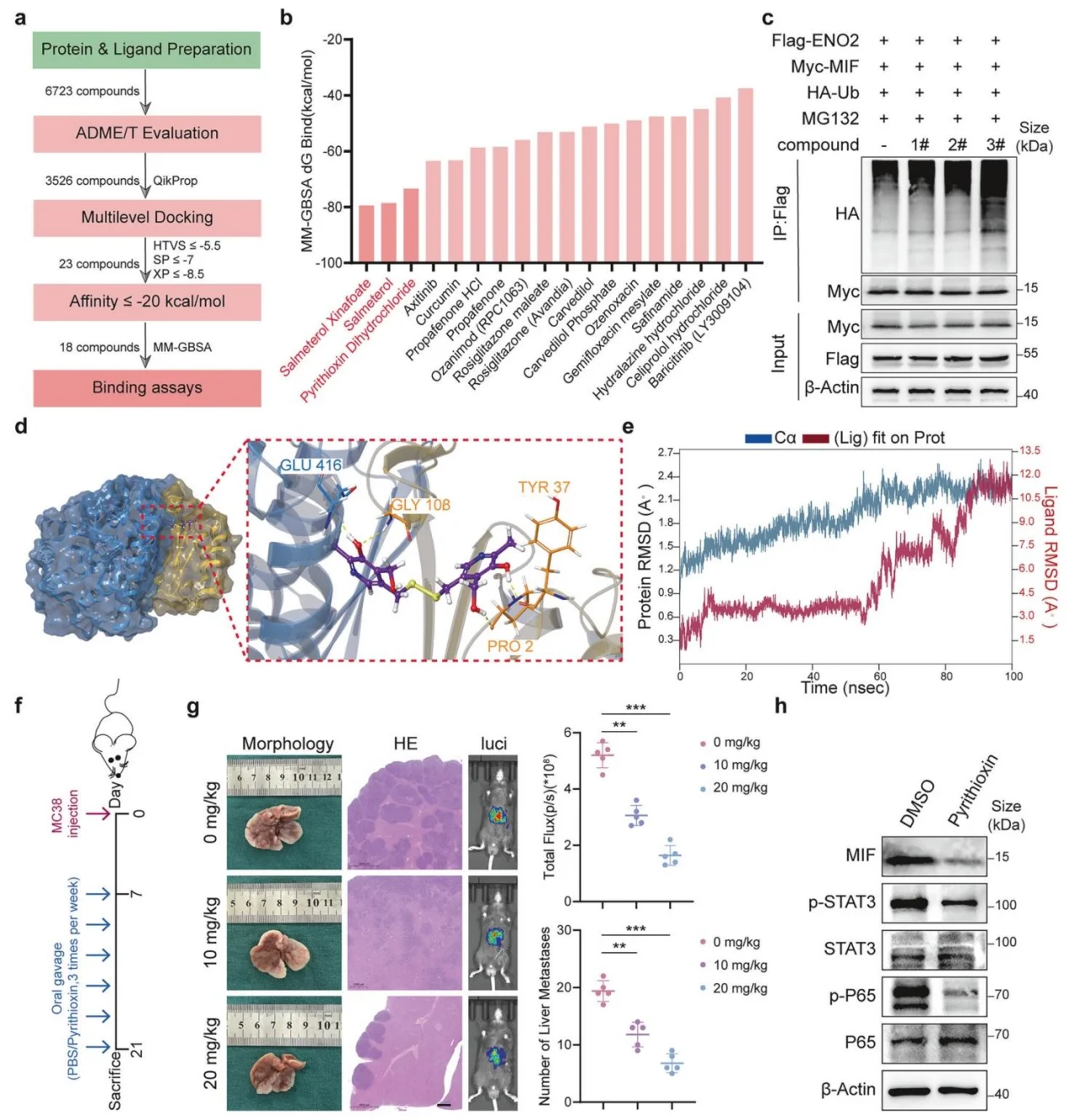
图8.吡硫氧辛作为ENO2-MIF相互作用抑制剂的鉴定与验证及其体内抗转移作用
研究意义与创新点
- 理论创新:首次定义ENO2-MIF为结直肠癌肝转移全新调控轴,揭示糖酵解酶ENO2非代谢新功能——通过蛋白互作重塑肿瘤免疫微环境;
- 机制创新:阐明ENO2招募HSP90、拮抗CHIP介导MIF泛素化降解的分子新机制,解析肿瘤细胞调控巨噬细胞M2极化的全新信号通路;
- 技术创新:结合单细胞+空间转录组,精准定位ENO2⁺肿瘤细胞与M2巨噬细胞的空间互作关系;
- 转化创新:发现老药吡硫醇可靶向ENO2-MIF互作,显著抑制肝转移,为CRLM尤其是MSS型难治患者提供低成本、可快速转化的靶向免疫新策略。
文章小结
本研究利用单细胞与空间转录组,筛选并证实ENO2是结直肠癌肝转移的关键驱动因子;ENO2通过直接结合MIF、招募HSP90,抑制CHIP介导的泛素化降解,稳定MIF蛋白并激活STAT3/NF-κB通路,进而诱导巨噬细胞M2极化、构建免疫抑制转移微环境,促进结直肠癌肝转移。靶向ENO2-MIF互作的吡硫醇可有效抑制体内肝转移,为CRLM机制研究、预后标志物开发及临床靶向治疗提供了重要理论与实验依据。
原文链接:https://doi.org/10.1038/s41392-026-02732-2
源井生物基因编辑服务与产品
源井生物一直秉持“让基因编辑更简单”的核心理念,持续迭代产品与服务,已累计实现13000+基因编辑成功案例,拥有10000+ KO细胞产品,基于自主研发的创新技术,基因编辑效率相比于传统方法提升10-20倍。目前源井生物已为超10000个生命科学实验室、药企和CRO公司提供优质的基因编辑服务与产品。
ENO2基因,也称为NSE,是Enolases家族的一员。这个家族的基因主要编码烯醇化酶,这是一种在糖酵解过程中起关键作用的酶。ENO2基因不仅在糖酵解中起作用,还参与调控细胞内的信号通路。如果您也想针对ENO2展开研究,源井生物可为您提供ENO2基因敲除细胞,涵盖HCT-116、HCT15、293T细胞系,还有更多热门靶点KO细胞,保证WB!如果您有个性化基因编辑的需要,欢迎咨询!







