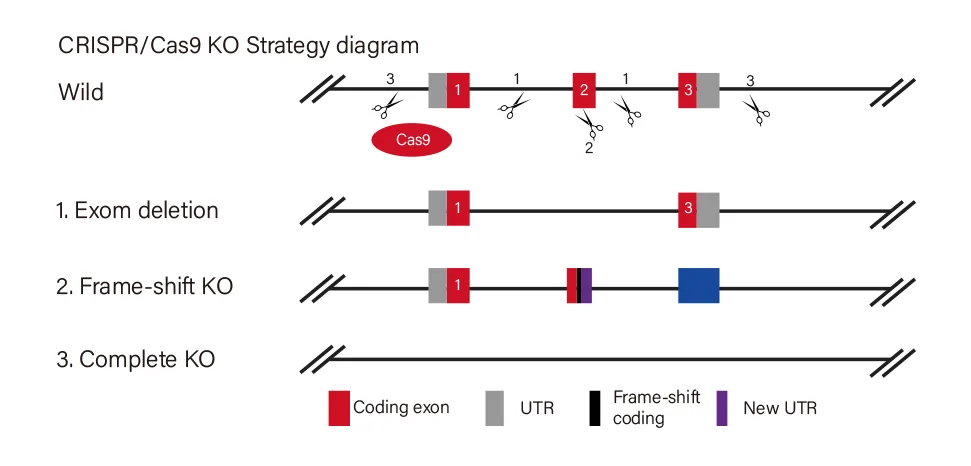
EZ-editor™技术平台
EZ-editor™是源井生物针对细胞系基因编辑自主研发的技术(基于CRISPR/Cas),适用于高效基因敲除细胞的构建。该技术整合了多项核心能力,包括:
相较于传统方法,源井生物EZ-editor™技术可将细胞基因编辑效率提升10-20倍。
基因敲除细胞系服务详情
编辑类型
单基因敲除/多基因敲除
细胞类型
肿瘤细胞、永生化细胞系、iPS以及各类难转染的细胞
价格(CNY)
询价
项目周期
快至4周
交付标准
1.纯合敲除单克隆细胞株 (>106个细胞/管,每个克隆各2管); 2.野生型细胞 (>106个细胞/管,1管); 3.实验报告;
敲除验证
PCR检测(常规)、Sanger测序(常规)、Western Blot(可根据实验需求评估)
其他质控
通过STR鉴定,细菌、真菌、支原体检测;通过细胞复苏活率检测;

源井基因敲除细胞服务优势
500+基因编辑成功案例
消化系统

内分泌系统

呼吸系统

生殖系统

循环系统

血液,淋巴系统

脑和神经系统

泌尿系统

骨、关节、软组织、皮肤系统

干细胞

眼耳鼻喉口腔系统

基因敲除细胞方案
源井生物提供三类标准敲除方案,并可根据客户科研目标进行定制化调整
服务流程及方案适配
项目评估&红棉设计敲除方案
了解研究目标,推荐最合适的
敲除策略与细胞类型

RNP复合物
合成sgRNA,准备Cas9蛋白
细胞转染(电转)
采用电转法(Nucleofector等平台)
将RNP导入细胞,优化电转程序以最
大化存活率与编辑效率

Cell Pool效率验证
检测Cell Pool的转染效率和切割效率

单克隆筛选
单细胞扩增,挑选足够的阳性克隆
进行测序验证

阳性验证&质量检测
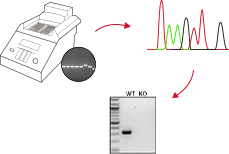

成果交付



精选案例分享
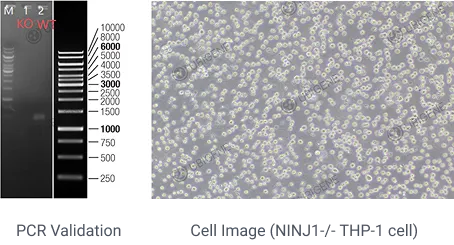
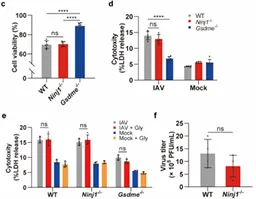
1. Ninjurin-1 mediates cell lysis and detrimental inflammation of PANoptosis during influenza A virus infection
IF=52.7
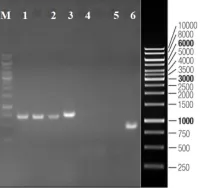
2. Activation of lysophagy by a TBK1-SCFFBXO3 TMEM192-TAX1BP1 axis in response to lysosomal damage
IF=15.7
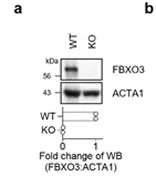
引用文献节选
Hep G2细胞敲除PPP1R12B/KCNJ12/FGA基因——肝细胞癌
IF=64.8
Nature
Deep whole-genome analysis of 494 hepatocellular carcinomas
该研究完成了中国人群肝细胞癌全基因组深度特征分析,还选取了3个新鉴定的潜在驱动事件进行详细功能验证,发现上述基因的突变足以导致基因表达水平的显著变化(其中PPP1R12B,KCNJ12,FGA基因敲除细胞和携带突变位点的过表达慢病毒均由源井生物构建),并参与调控肝细胞癌的多种恶性表型,这些结果证实基于数据分析发现的新驱动事件的有效性。 查看详情>>
HeLa细胞系敲除STING1基因——病毒防治
IF=32.4
Immunity
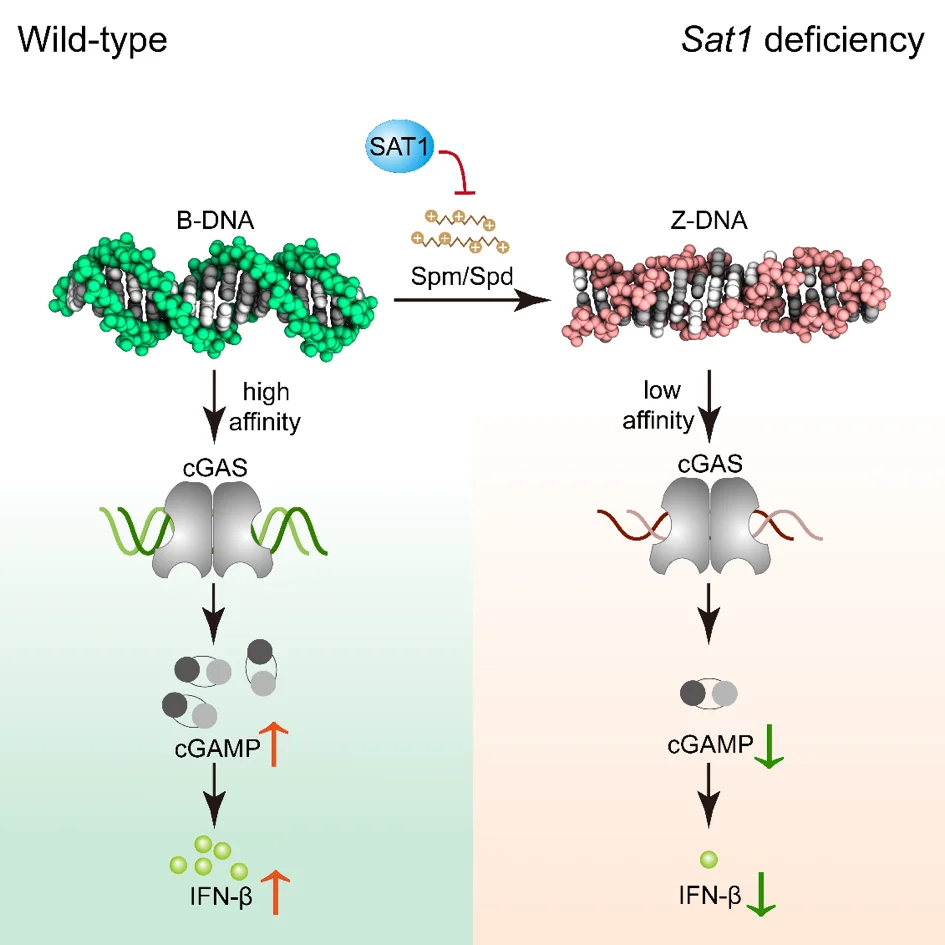
Polyamine metabolism controls B-to-Z DNA transition to orchestrate DNA sensor cGAS activity
为了深入研究cGAS活性调节机制对于预防和治疗病毒性疾病的作用,山东大学赵伟教授课题组采用源井生物构建的敲除STING1基因的HeLa细胞 系作为关键模型,发现DNA的两种构象(B-DNA及Z-DNA)与cGAS亲和力不同;内源性代谢小分子精胺和亚精胺及多胺分解代谢关键酶SAT1通过诱导DNA构象转换调节cGAS活性。该研究揭示了一种防止细胞DNA异常识别的新机制,并为治疗cGAS活性异常相关疾病提供了治疗靶点。 查看详情>>
HCT116细胞系敲除TP53基因——细胞凋亡
IF=16.6
Nature Communications
Structures of p53/BCL-2 complex suggest a mechanism for p53 to antagonize BCL-2 activity
该研究采用源井生物构建的敲除TP53基因的HCT116细胞系,通过解析p53与抗凋亡蛋白BCL-2的复合物晶体结构,并结合生化与细胞实验,揭示了p53与BCL-2蛋白相互作用并促进细胞凋亡的新机制,即p53通过直接占据BCL-2的BH3结合口袋与之形成复合物,并通过释放位于口袋的促凋亡BCL-2家族蛋白来拮抗BCL-2活性,从而促进细胞凋亡 。这些结构和功能数据为进一步了解p53介导的线粒体凋亡的复杂调控机制提供了新的思路,也为研发靶向蛋白质间相互作用激活细胞凋亡的抗癌治疗策略提供了重要基础。 查看详情>>
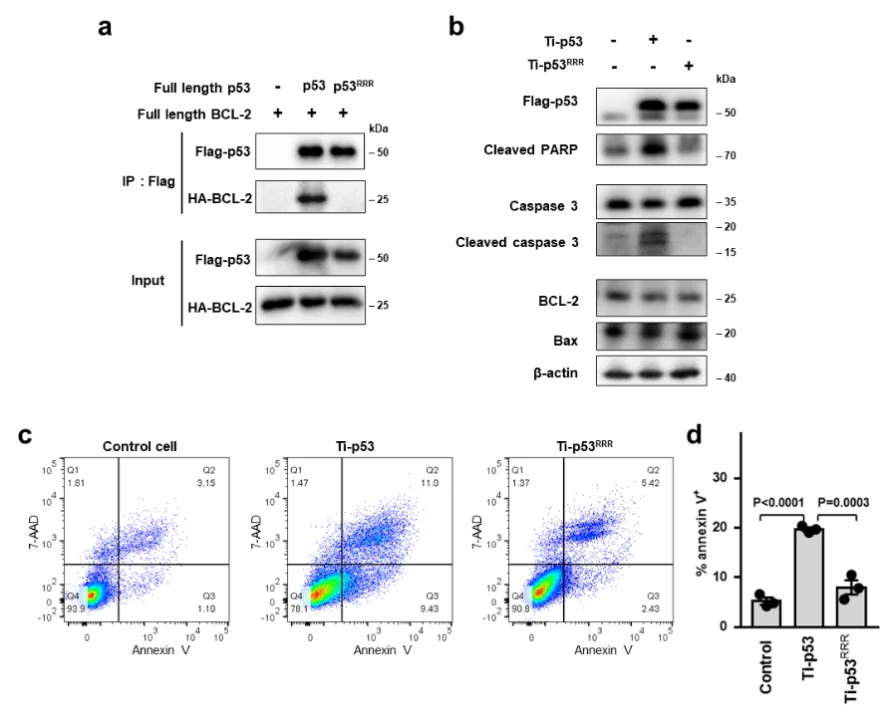
U2OS细胞敲除ZNF432基因——卵巢癌耐药
IF=14.9
Nucleic Acids Research
ZNF432 stimulates PARylation and inhibits DNA resection to balance PARPi sensitivity and resistance
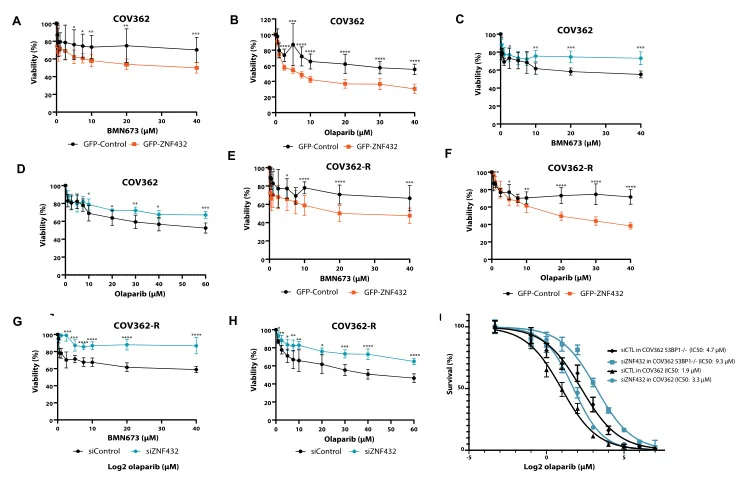
ZNF蛋白被证实为调节哺乳动物细胞基因组完整性的关键因子,为探究ZFP可作为基于同源重组(HR)的DNA修复效应器的可能性,拉瓦尔大学Jean-Yves Masson 团队采用源井构建的U2OS敲除ZNF432细胞系进行了一系列实验,发现ZNF432在癌细胞中缺失会加速DNA修复,导致PARPi作用减弱,使卵巢癌细胞产生耐药性,证实ZNF432是一种新的HR抑制因子,成功拓宽了研究PARPi疗效的新途径。 查看详情>>
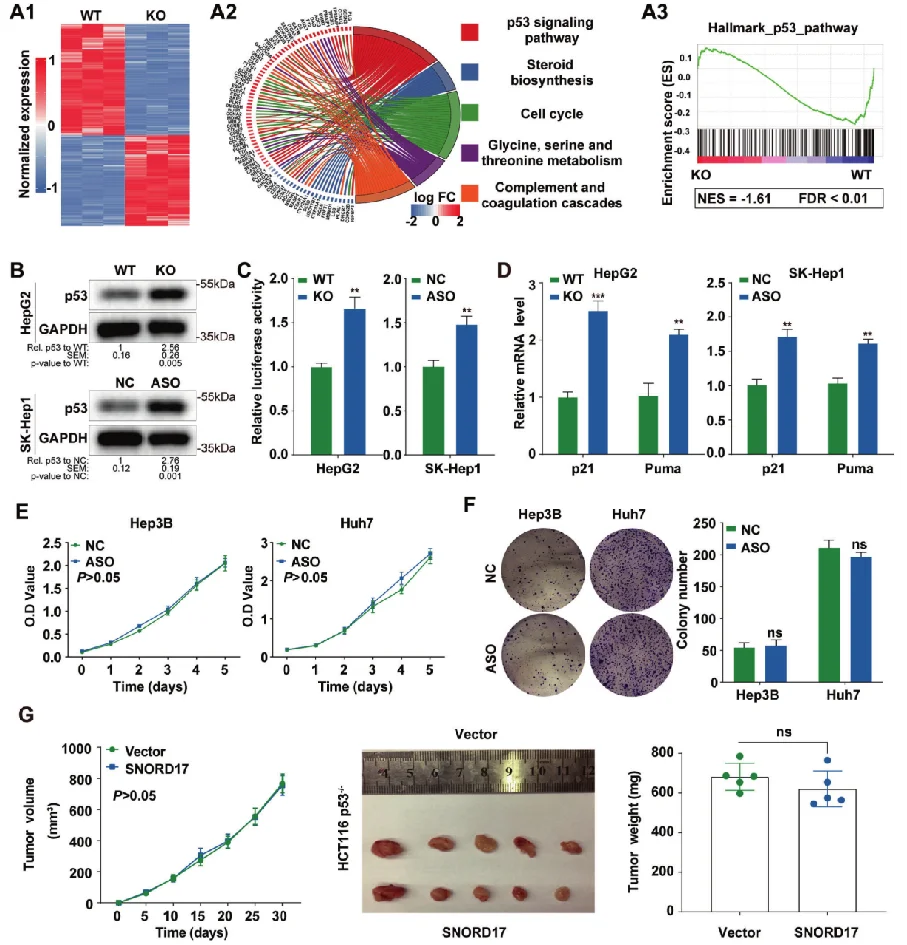
Hep G2细胞敲除SNORD17基因——肝细胞癌
IF=12.4
Cell death & differentiation
Non-coding small nucleolar RNA SNORD17 promotes the progression of hepatocellular carcinoma through a positive feedback loop upon p53 inactivation
该研究构揭示了小核仁RNA SNORD17和p53信号通路在肝细胞癌的中的调控作用,为肝细胞癌的治疗提供了一个新的潜在靶点 。源井生物为本研究提供关键细胞模型:SNORD17基因敲除的Hep G2细胞,通过体外和体内实验表明,在HCC细胞系敲除SNORD17基因可显著抑制细胞增殖、克隆形成和G1/S期转变。 查看详情>>
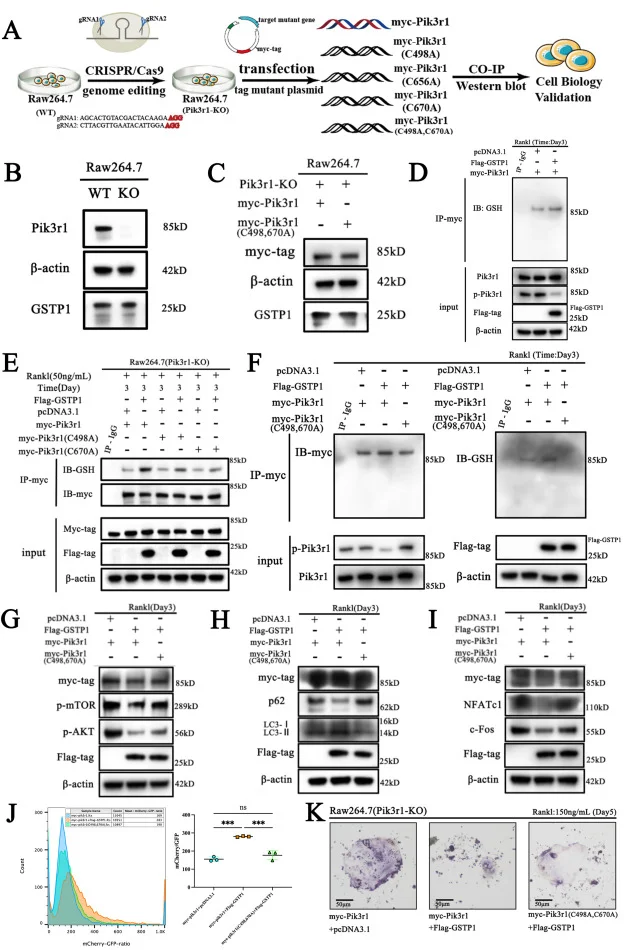
RAW264.7细胞敲除Pik3r1——骨质疏松
IF=11.4
Redox Biology
GSTP1-mediated S-glutathionylation of Pik3r1 is a redox hub that inhibits osteoclastogenesis through regulating autophagic flux
该文章通过源井生物构建的Pik3r1基因RAW264.7细胞敲除模型 ,结合大量的体内外实验探索了GSTP1重塑破骨细胞相关骨稳态的一种全新机制,第一次诠释了破骨细胞的细胞命运是由GSTP1介导的氧化还原相关的翻译后修饰-自噬流级联轴来决定的。 查看详情>>
FAQs
1. 什么是基因敲除系?
基因敲除细胞系(Gene Knockout Cell Line) 是指在特定细胞中 人为删除或失活某个基因 ,从而使该细胞 不再表达该基因编码的蛋白质 的细胞系。这种细胞系常用于 研究该基因的功能 ,以及它在细胞生理、生化途径、疾病机制中的作用。常用的方法如CRISPR-Cas9技术:利用Cas9核酸酶在目标基因处产生双链断裂,随后通过非同源末端连接(NHEJ)导致框移突变,使基因失活。
EZ-editor™ 是源井生物自主研发的高效细胞基因编辑平台,基于 CRISPR/Cas9 技术,结合独特的 gRNA 设计算法、丰富的细胞系编辑参数数据库、精确的 Pool 编辑效率检测方法、单克隆形成率提升策略,以及适用于微量细胞的高通量基因型鉴定体系。在基因敲除细胞系构建方面,源井生物自主研发的 EZ-editor™ 技术相较于传统方法, 可将基因编辑效率提升 10–20 倍。
2. 基因敲除与敲低有什么区别?
基因敲除细胞系是通过CRISPR/Cas9等手段 永久性删除目标基因, 可以实现使其失活;而基因敲低细胞系则通过siRNA或shRNA等方式 暂时性或部分抑制基因表达 ,基因本身仍存在。敲除适用于深入研究基因功能或构建疾病模型,敲低则更适合初步筛选或研究必需基因。两者在稳定性、效率和应用场景上各有优势。
源井生物不仅提供基因敲除细胞系构建服务,还可为您定制高效的 基因敲低(RNA干扰) 解决方案。无论您的研究目标是基因功能验证、机制探索,还是疾病模型构建,源井生物都能为您提供可靠、高效的基因编辑解决方案。
3. 是否对KO细胞系进行了支原体检测?
我们所有KO细胞系在交付前均通过支原体检测,使用 源井生物EZ-Detect™支原体检查试剂盒 对75种常见及罕见支原体 检出结果均为阴性,并可以提供支原体检测报告。
4. KO细胞系质检标准是什么,有哪些承诺?
源井生物提供的KO细胞系质检涵盖PCR检测、Sanger测序、支原体及菌检、STR鉴定等多项内容,从多个层面保障细胞质量与来源可靠性。源并生物所有交付的KO细胞均已通过PCR检测与Sanger测序,确保KO细胞在基因组水平上的敲除效果。此外,根据客户的具体实验需求,还可增加Western Blot检测,以进一步在蛋白水平验证基因敲除效果。
5. 我可以使用自己的野生型细胞系吗,还是必须购买你们的细胞系?
客户可以自己提供细胞系,若客户没有细胞,公司可免费提供细胞系,源井生物拥有 超1000种野生型细胞 ,含大量基因编辑成功案例细胞,可用于各类基因编辑实验;也可根据客户需求,从细胞库采购特定细胞系,完成KO细胞系构建后交付。
若客户自己提供细胞系,客户提供的细胞系需满足以下条件:
- 细胞类型明确,活力≥95%,汇合度70%-80%,无细菌、真菌、支原体污染,可以长期增殖和传代,状态良好;
- 提供细胞培养说明书(含培养基、血清、培养条件等),便于我们后续培养和转染;
- 细胞数量:3瓶活细胞,T25/瓶;或者2管冻存细胞,1*10^6/管,确保有足够的细胞进行转染和筛选。
6. 你们的细胞生物安全等级(BSL)是几级?
我们提供的KO细胞系通常在 BSL-1或BSL-2 条件下进行培养和质检,并严格遵循相关生物安全管理规范。如需针对具体细胞系的BSL等级信息,可联系客服进行确认。
7. 应该如何培养KO细胞呢?
培养步骤参考细胞说明书,推荐使用我们的 KO细胞专用培养基 进行培养,可有效恢复并稳定基因编辑后细胞的生长状态,提升细胞存活率与功能性表现,确保基因敲除细胞株在长期培养中保持稳定。我们也可以提供 专业的技术支持 ,全程指导KO细胞培养。
8. 基因敲除服务多少钱?
我们的定制化基因敲除服务使用自主研发的EZ-editor™基因编辑技术,相比于传统方式编辑效率提升10-20倍, 提前4周 筛到阳性克隆。敲除细胞系服务提供PCR检测及Sanger测序验证,确保基因编辑的准确性。如有特殊验证需求(如Western Blot蛋白水平验证),可根据具体项目进行评估并提供相应服务。 点击在线咨询 ,获取技术评估方案和报价。
此外,我们还提供现货KO细胞,截至目前源井生物已构建 10000+KO细胞产品 ,承诺WB!低至¥5980 !最快1周交付。









